Skin - Integumentary System
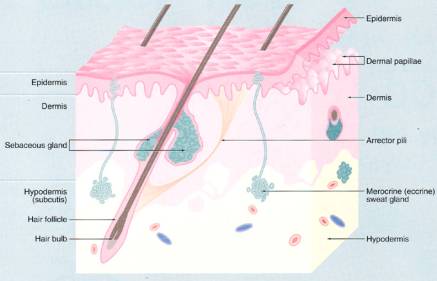
The integumentary system (integument, a covering) consists of the skin (cutis) and its appendages. The protective covering for the body known as the skin is one of the largest organs of the body and accounts for approximately 16% of the body weight. It contains glands, blood vessels, lymphatic vessels, nerves, and smooth muscle (arrector pili muscles). The appendages are derived from the overlying epidermis and include two types of sweat glands, sebaceous glands, hair, and nails.
The skin has two defining tissue types, epidermis (keratinocytes are derived from ectoderm) and dermis (derived from mesoderm). The dermis interfaces with the underlying hypodermis, which is the fatty subcutaneous connective tissue or the superficial fascia of gross anatomy. Although the hypodermis not officially considered to be one of the layers of the skin, it is a loose connective tissues that attaches the skin to underlying tissues by way of vertical septae and permits skin mobility over most parts of the body (e.g., when it is pinched up), it serves as an insulator and shock absorber and serves as a caloric reservoir.
Skin is often classified as either thick skin or thin skin. This classification refers to epidermal thickness rather than dermal thickness or to the thickness of the skin as a whole. Thick skin (= thick epidermis) is restricted to palms and soles. Thin skin (= thin epidermis) covers the remainder of the body. Although skin may have a very thick dermis in some sites, as on the back, it is classified as thin skin if the epidermis is thin.
Skin has many functions, some of which can be appreciated from the study of its microscopic structure. As the covering of the body, the skin provides some protection from injury, and from the entry of microorganisms. Protection is also provided by the melanocytes, which produce pigment to shield the body from too much ultraviolet light. The skin functions in temperature regulation, e.g., by sweating, varying peripheral blood flow. It covers the surface with sebum to lubricate the epidermal surface and prevent cracking. The adipose tissue of the hypodermis provides insulation. The skin also functions in excretion (and secretion) via its sweat and sebaceous glands. Moreover, it stores fat, manufactures vitamin D and is the first guard of the immunosurveillance system. Cutaneous receptors for touch, pressure, heat, and pain stimuli provide a sensory appreciation of the environment; thus the skin serves as a sensory organ. The skin acts as a friction surface for motor tasks involving grasping, rubbing, scratching, etc. The skin is also important in preventing the loss of body fluids, a fact that is well illustrated when excessive fluid loss occurs after extensive burns.
Thick Epidermis/Thick Skin
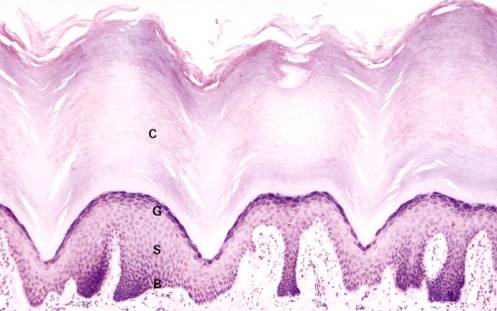
The predominant cell type, an epidermal keratinocyte begins its life cycle in the basal layer and becomes progressively more differentiated as it moves toward the surface - a process that produces a stratified squamous keratinizing epithelium.
Observe the flattened, dead surface cells (sometimes called corneocytes or dandruff in the scalp) of the thick stratum corneum. A few of the surface cells are being desquamated. Some areas of the stratum corneum show spiraling sweat ducts, each of which will end in a sweat pore at the surface. Nuclei and other cellular organelles are absent in the stratum corneum.
The stratum granulosum is composed of increasingly flattened rectangular-shaped cells filled with keratohyalin granules. The large accumulation of basophilic granules in these cells is composed of filaggrin and keratin filaments. The thickness of this layer varies from one to three cells and as a rule is representative of the rate of keratinization. Thus, there are little or no stratum granulosum or keratohyalin granules in diseases, with a rapid rate of keratinization such as psoriasis, whereas the granular layer is thickened in diseases with a slow rate of keratinization.
The stratum spinosum is composed of several layers of polyhedral cells, which flatten as the stratum granulosum is approached. See Figure left. The cytoplasm is rich in tonofibrils, which are specific for keratin pairs 1 and 10. Individual cells appear to be separated by spaces that are traversed by the so-called intercellular bridges which, under oil immersion, can be seen as fine lines between adjacent cells. 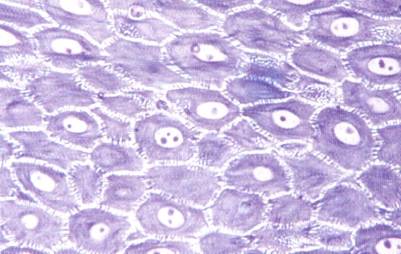
They account for the prickly appearance of the cells and for their frequent designation as prickle cells. These are tissue preparation artifacts caused when water is removed during the dehydration step. Spinous cells are attached to their neighbors by a multitude of desmosomal attachments and these connections remain as the fragile cytoplasm shrinks.
The stratum basalis is composed of a single layer of cuboidal to columnar cells, which are attached to the basement membrane by hemidesmosomes. These cells express keratin intermediate filament pairs 5 and 14. This layer contains the stem cell population and an occasional mitotic figure of a transient amplifying cell may be observed. A few melanocytes (1 for every 36 keratinocytes) can be identified by their pale (empty) cytoplasm and dark nuclei among the keratinocytes of the stratum basalis. These cells do not contain tough keratin filaments and also shrink during tissue preparation.
A basement membrane exists between the epidermis and dermis. Reticular fibers are present in the basement membrane and account for the prominence of this structure after PAS staining. Note that the border between the epidermis and dermis known as the dermal-epidermal junction is irregular because numerous upward interdigitations known as dermal papillae indent the undersurface of the epidermis. Papillae are far more prominent in thick skin than in thin skin since they serve to help these surfaces resist shear forces and friction. Rete pegs are corresponding elongated downward projecting ridges of epidermis. Some rete pegs are narrower than others are and they are the ones, which receive the sweat ducts. Under oil immersion, note that the basal cells of the larger pegs have very fine processes, which anchor the cells to the basement membrane. Basal cells of the narrow ridges are smoother, allowing them to shift more easily.
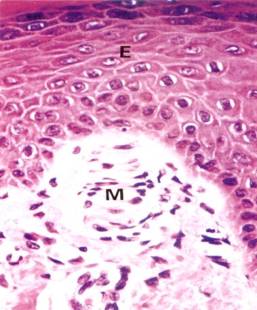
Numerous capillaries are present in the papillae of the papillary layer. Meissner's corpuscles (encapsulated sensory nerve endings for touch that resemble sectioned pinecones) can be seen in some papillae (see Figure below). The reticular layer of the dermis is so named because of the woven arrangement of the heavily cross-linked collagenous fibers (in the form of a close meshed net) and not because of the presence of reticular fibers. Note that the collagenous fibers, although cut in transverse, oblique, and longitudinal planes, run parallel and not vertical to the epidermis.
Secretory portions of eccrine sweat glands can be seen deep in the reticular dermis and extending deeper into the upper hypodermis. In addition to the secretory portions of sweat glands, the hypodermis (subcutaneous connective tissue) contains lobules of adipose tissue, which are separated by strands of collagenous fibers. When these septae become excessively filled with well-nourished adipocytes, the surface of the skin becomes dimpled and the appearance is known as cellulite.
Thin skin
The epidermis of thin skin has far fewer cells in each of its layers (i.e., the total thickness of the stratum corneum, stratum granulosum, and stratum spinosum are greatly reduced in thickness). Identify the melanocytes with dark nuclei and pale cytoplasm that can be seen interspersed between the keratinocytes in the stratum basale. Note that the brown melanin pigment that is present is confined to the keratinocytes in either the basal epidermis or those in the hair bulb. A few pigment containing macrophages can occasionally be seen in the dermis. These cells are phagocytic and do not produce pigment. Note that the dermal papillae and rete pegs are less developed than in thick skin thus the waviness of the dermal-epidermal junction is not as pronounced. The dermis is comparable to that of thick skin in that it has a thinner, more superficial papillary layer and a thicker, more deeply situated reticular layer.
The small blood vessels of the subpapillary plexus are located at the junction of the reticular layer and the papillary layer. Vessels arising from this plexus can be seen as capillaries and arterioles in the dermal papillae. The papillary blood vessels are in close association with the epidermal cells, which require nourishment for their growth and development.
Melanin, the pigment largely responsible for skin color, is produced by melanocytes, which number about the same in all races but differ in the amount of melanin they produce. Melanin is transferred via long cytoplasmic processes of melanocytes to the intercellular spaces where it is taken up by keratinocytes and placed in a supranuclear position melanosomes. Cell bodies and cytoplasmic processes of melanocytes are not clearly shown by conventional stains, but they can be demonstrated by treatment of sections with dihydroxyphenylalanine (DOPA). The DOPA reaction demonstrates that the cell bodies of most melanocytes are located in the stratum basale while their cytoplasmic processes (some times called dendritic processes) extend between cells of the stratum spinosum. Cells from the melanocytic lineage can also be identified by immunostaining using an S-100 antisera.
Epidermal Appendages/Adnexal Structures
The small, cuboidal cells -bordering the basement membrane undergo mitosis to replenish the more central ones which accumulate lipid and are lost as the glandular secretion. As a cell accumulates fat droplets, it enlarges to become polyhedral or spherical in shape. The nucleus becomes pyknotic and the entire cell eventually disintegrates to enter the sebaceous gland duct as sebum. This mode of secretion in which the entire cell is lost is called holocrine secretion, and is characteristic of sebaceous glands. Find a duct, which connects with the alveoli of a sebaceous gland. Ducts are lined with stratified squamous epithelium and open into hair follicles. Sebaceous glands in some locations (e.g., those in the margins of the lips, nipple, and glans and prepuce of penis) do not connect with hair follicles, but directly onto the surface of the skin.
There are two classes of sweat glands (sudoriparous glands), merocrine and apocrine. Most sweat glands of the body are of the merocrine type and are usually referred to as eccrine sweat glands. They are most numerous in the skin of the palms, soles, and forehead. It has been estimated that they number about 3000 per square inch in the palm of the hand. They are called merocrine glands because they secrete by the merocrine method, i.e., no part of the cell is lost with the secretion. Secretion leaves the cells in a manner that might be described as reverse pinocytosis. Apocrine sweat glands are encountered in only a few areas, e.g., in the axillae and in the anogenital region. They are called apocrine glands because they secrete by the apocrine method, i.e., the apical portion of the cell breaks off to form part of the secretion. Both types of sweat glands are simple coiled tubular glands. This means that each sweat gland has its own duct (one duct per gland) and that the secretory portion of the gland is a small-coiled tube.
Secretory portions of the glands lie deep in the dermis, at the dermal-hypodermal junction, or occasionally within the hypodermis. A section through the secretory portion of a sweat gland will cut the coiled tubule into several cross sections (see Figure left photo). It may appear that two layers of secretory cells line a tubule, but the outermost cells with somewhat elongated nuclei are the myoepithelial cells, which occupy a constant position between the basement membrane and the base of the secretory cells (see Figure right photo). Myoepithelial cells have long contractile processes, which embrace the secretory cells and squeeze out the secretion. The processes are not visible in H&E preparation, but they can be demonstrated by the alkaline phosphatase technique. Myoepithelial cells are also present in other glands, salivary glands, and mammary glands, and in each instance they occupy a similar position between the basement membrane and the base of the glandular epithelial cell. In addition to basally located myoepithelial cells, pale cuboidal secretory cells can be seen surrounding the lumen of the secretory tubule.
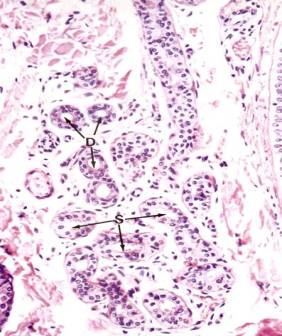
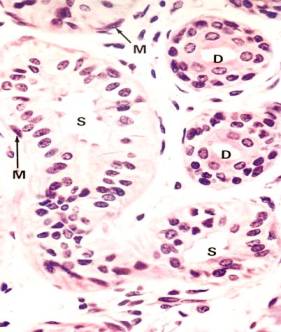
Intradermal sweat ducts are lined by two layers of cells whose cytoplasm is darker than that of secretory cells. The two-layered epithelium of the sweat duct is classified as stratified cuboidal. Myoepithelial cells are absent in the duct.
Secretory portions of apocrine glands are much larger than secretory portions of eccrine glands and their lumina measure as much as 200 μm in diameter (see Figure 61 below). This is ten times the average diameter of lumina of eccrine sweat glands. Apices of some cells of apocrine glands are rounded and can be seen breaking off to enter the lumina as part of the secretion. This mode of secretion where part of the cell is lost is termed apocrine secretion. Myoepithelial cells are present in secretory portions of apocrine glands, and appear even more prominent than in eccrine glands.
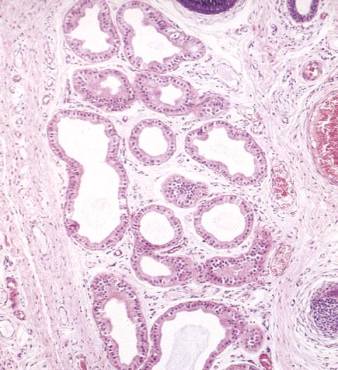 Micrograph of apocrine sweat glands.
Micrograph of apocrine sweat glands.
The intradermal ducts of apocrine glands have the same histological appearance as corresponding ducts of eccrine glands. They are devoid of myoepithelium and have a double layer of dark staining epithelial cells, which have a periluminal cuticle. The latter becomes especially prominent as the duct traverses the papillary layer of the dermis. Most ducts of apocrine glands open into hair follicles, but some open directly onto the surface, as do all eccrine ducts. Quite commonly, more than one apocrine duct empties into the same hair follicle, usually entering the follicle slightly above the opening of the sebaceous duct.
Apocrine and eccrine glands have other differences in addition to those of histological importance. Eccrine glands are supplied with cholinergic fibers while apocrine glands are supplied with adrenergic fibers. Eccrine glands serve primarily in heat regulation whereas apocrine glands represent scent glands similar to those producing pheromones in lower animals. Both types of glands are stimulated to secrete by stresses such as fright and pain, but eccrine glands also respond to heat while apocrine glands do not. In contrast to eccrine glands, apocrine glands are greatly influenced by hormones, becoming active about the time of puberty and showing secretory variation during the menstrual cycle.
Hair Follicles/Nails
Keratinocytes that form these appendages grow downward from the surface during the 3-4 month of development. The stem cell population resides in the bulge region where the erector pili muscle inserts into the hair shaft. Periodically these cells grow downward (anagen growth phase), form an expanded hair bulb in the deep dermis or hypodermis and begin to undergo a special pattern of differentiation that will produce a highly keratinized structure known as hair. The transiently amplifying hair matrix cells are found at the upper indentation of the hair bulb above of a tuft of capillaries, the dermal hair papilla that provide nourishment. Melanocytes also populate the hair bulb region and provide coloration to the hair. The cells lining the outer surface of the length of the hair follicle are known as outer root sheath cells and the multiple inner cell layers are known as inner root sheath cells.
Hairs (terminally differentiated cells that are equivalent to dead stratum corneum squames on the surface) move upward where they become coated with sebum (oil) and eventually erupt at the epidermal surface. Hair (totally dead cells) has an outer cuticle and an inner cortex. Periodically the hair bulb begins to regress upward to the bulge region (catagen phase), the hair is shed at the surface, and the hair follicle enters the telogen (resting) phase. The length of the hair growth cycle and the type of hair (terminal hair on the head, vellus-fine hair, pubic hair) is determined by local signals as well as hormones.
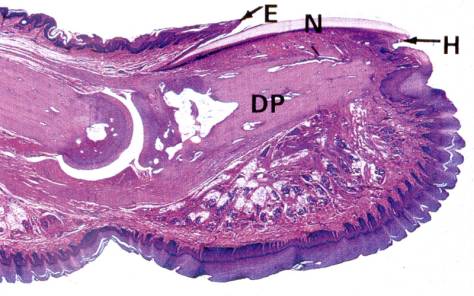 Micrograph of a developing fingernail on the distal phalanx (DP). E = eponychium, N = Nail, H = hyponychium.
Micrograph of a developing fingernail on the distal phalanx (DP). E = eponychium, N = Nail, H = hyponychium.
Fingernails and toe nails represent yet another type of differentiation (see Figure). The nail matrix cells are located under the skin fold at the proximal region of the distal phalanx. Destruction of nail matrix cells will result in a permanent nail loss. The more differentiated cells advance toward the fingertip. The newly differentiated cells form a crescent-shaped lunula (white region that is grossly visible). The proximal dead skin that is carried forward by continuous nail growth is known as the eponychium and the fold of skin underneath the distal growing tip is called the hyponychium. The dead nail advances across epidermal cells called the nail bed.
Advertise With Us
Latest Health News
- WHO urges cost effective solutions on NCDs and mental health amidst slowing progress - World Health Organization (WHO)
- Seven ways your coffee habit could be ruining your health, from digestion to cholesterol - The Telegraph
- Six Tasmanian aged care facilities share in $31 million upgrades - Australian Government Department of Health, Disability and Ageing
- Healthcare provider blindsided by huge change at cancer treatment centre - Australian Broadcasting Corporation
- ‘Blanket exclusion’: life insurers block mental health conditions - The Australian